Axe 1 : Évolution des asymétries de l'habenula chez les vertébrés
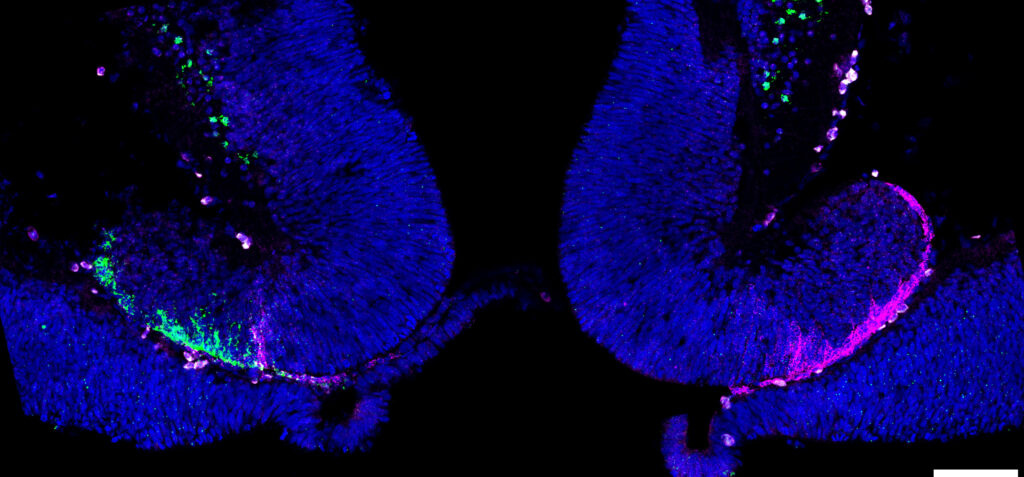
La diversification des identités neuronales au cours de l’ontogenèse et de l’évolution dans le cerveau antérieur des vertébrés est une question fascinante. Les habenulae, structures épithalamiques bilatérales connues pour leur rôle dans le traitement des stimuli aversifs, offrent un excellent système pour aborder cette problématique. Chez de nombreux vertébrés, elles présentent des asymétries qui peuvent considérablement varier en nature et en degré entre le côté gauche et le côté droit. Notre travail vise à déchiffrer les tendances et les forces motrices qui façonnent ces variations chez les vertébrés. Pour ce faire, nous nous concentrons principalement sur deux espèces sélectionnées pour leur position phylogénétique et les asymétries marquées de l’habenula qu’elles présentent : un poisson cartilagineux, la petite roussette Scyliorhinus canicula, et un cyclostome, la lamproie Lampetra fluviatilis. Les résultats obtenus chez la petite roussette indiquent que l’évolution des habenulae a suivi des voies différentielles chez les actinoptérygiens et les sarcoptérygiens, à partir d’une organisation ancestrale hautement asymétrique conservée par les chondrichthyens. Ils suggèrent également que cette diversification a impliqué une logique développementale conservée, reposant sur des interactions entre une régulation temporelle de la neurogenèse et une activité asymétrique de Wnt, modifiant les identités neuronales et sujette à des variations évolutives dans le temps et l’espace. Notre travail vise à tester davantage ces hypothèses via des analyses d’un plus large éventail de vertébrés et la combinaison de profilages 3D et d’approches fonctionnelles. Nous nous concentrons sur trois questions principales :
1. Explorer les tendances évolutives des asymétries de l'habenula dans les principaux taxons de vertébrés.
Cette analyse vise à obtenir des caractérisations non biaisées des asymétries de l’habenula chez un large éventail de vertébrés (par exemple, lamproie, petite roussette, dipneuste, bichir et chromis épineux), en utilisant des combinaisons d’approches snRNA-seq et de profilage ARN 3D (Tomo-seq, Stereo-seq). L’analyse des schémas de projections efférentes est menée en parallèle, en collaboration avec le groupe de M. Concha, au Chili.
2. Générer une référence chondrichthyen pour l'élaboration des asymétries au cours du développement.
Avec son riche répertoire d’asymétries de l’habenula, dont certaines reflètent probablement des traits ancestraux, la petite roussette est un excellent modèle pour cette question. Nos analyses visent à établir des trajectoires transcriptionnelles cellulaires dans l’habenula en développement de la petite roussette en utilisant snRNA-seq et HCR-FISH. Nous abordons également les mécanismes régulant les identités et les asymétries des progéniteurs, en mettant l’accent sur la signalisation Wnt.
3. Diversifications mécanistiques chez les cyclostomes et les téléostéens.
Cette analyse se concentre sur trois espèces (lamproie, chromis épineux, poisson-zèbre), en utilisant la petite roussette comme référence (voir l’axe 2 ci-dessus). Elle pourrait permettre la génération de larves/juvéniles présentant des schémas d’asymétrie perturbés et pouvant être soumis à des tests comportementaux chez la lamproie et le chromis épineux (en collaboration avec Marc Besson, Laurence Besseau et Michaël Fuentes, BIOM). De tels outils ouvriraient de nouvelles perspectives pour aborder la signification biologique de la diversification des asymétries de l’habenula, une question clé encore sans réponse.
Axe 2 : Diversifications du mode de formation du mésoderme chez les vertébrés
Les résultats obtenus dans le cadre d’un projet collaboratif avec R. Reshef (Université de Haïfa) et M. Schubert (Observatoire Océanologique, Villefranche-sur-Mer) ont soulevé la question d’une contribution ancestrale du mésoderme somitique segmenté au mésoderme intermédiaire pronéphrique (MI), traditionnellement considéré comme une composante mésodermique distincte et non segmentée, chez les vertébrés. Une question connexe a été soulevée en parallèle par l’équipe dirigée par S. Bertrand, qui a découvert que chez l’amphioxus, tous les dérivés du mésoderme non axial proviennent d’une couche mésodermique segmentée, contrairement aux vertébrés qui possèdent une composante mésodermique latérale non segmentée (mésoderme de la plaque latérale ou MPL). Ces observations ont conduit à un projet collaboratif conjoint avec ces groupes, visant à évaluer la contribution des somites au MI ou au MPL chez la lamproie et la petite roussette.
La stratégie expérimentale repose sur une combinaison de caractérisations morphologiques de la formation du mésoderme et d’approches RNA-seq. Nous utilisons l’imagerie 3D haute résolution et l’analyse de sections histologiques pour caractériser les composants mésodermiques MI et MPL et rechercher des preuves de segmentation lors de leur individualisation. Nous avons également développé des séries RNA-seq du développement précoce de la lamproie et de la petite roussette comme outil pour aborder le timing d’expression des marqueurs candidats du MPL (ou MI). Nous adaptons actuellement des approches snRNA-seq au développement précoce de la petite roussette et de la lamproie, afin de reconstruire les trajectoires transcriptionnelles sous-jacentes à la ségrégation des différents composants mésodermiques et d’inférer l’état ancestral des vertébrés.